REPRODUÇÃO
Os peixes têm uma extraordinária variedade de formas
de reprodução
Os peixes têm uma
extraordinária variedade de formas de reprodução, desde a partenogénese como em
Poecilia formosa, até aos machos
parasitas permanentemente fixos às fêmeas do peixe de profundidade Edriolychnus schmidti. da mesma forma, a fecundidade varia
entre os 300 milhões de ovos por ano em Mola
mola até uma descendência reduzida como em alguns tubarões. Os cuidados
parentais podem estar ausentes, como em muitos peixes pelágicos, ou envolver
diversas formas de vigilância do ninho ou eclosão bucal. Esta variedade implica
que a capacidade reprodutiva das populações de peixes também tenha formas
bastante diferentes.
Assim, o
conhecimento da reprodução é essencial para uma boa gestão e conservação das
espécies de peixes.
A
informação sobre reprodução na FishBase está estruturada em três tabelas:
REPRODUÇÃO; MATURAÇÃO e POSTURA.
A tabela
REPRODUÇÃO documenta a forma geral e tipo de reprodução que se aplica às
espécies em todas as suas formas.
As
tabelas MATURAÇÃO e POSTURA, por outro lado, apresentam informação sobre os
tamanhos e idades da primeira maturação e postura de diferentes populações da
mesma espécie em diferentes localidades. estas
tabelas são descritas a seguir.
Onde o óvulo e o espermatozóide se encontram
A tabela reprodução contém
informação sobre a forma de reprodução, frequência de postura, se existem
várias posturas ou não, e o padrão reprodutivo a que cada espécie corresponde.
A descrição do ciclo de vida, do comportamento, do acasalamento e postura,
também estão presentes nesta tabela.
Campos O modo de
reprodução está classificado nas seguintes escolhas: dioiecismo; protândrismo;
protogenia; verdadeiro hermafroditismo; partenogénese. O modo de Fertilização refere-se aonde o óvulo e
o espermatozóide se encontram, o que pode ser externo, interno (oviduto), na
boca, em bolsas externas ou estruturas similares, ou outros.
A
frequência da postura é descrita pelas seguintes escolhas:
·
Um pico sazonal por ano
(i.e., a época de postura é curta, com duração de apenas umas semanas ou meses
e pouca ou nenhuma postura ocorre fora dessa época)
·
Todo o ano, tendo um
pico (i.e., a postura ocorre durante todo o ano, mas existe um largo pico
sazonal).
·
Dois picos sazonais por
ano (i.e., a postura pode ocorrer durante todo o ano, mas dois picos são
claramente visíveis (geralmente um maior que o outro, separados por 5 a 7
meses)).
·
Sem nenhum pico sazonal
(i.e., a postura decorre todo o ano, sem nenhum pico sazonal bem definido)
·
Variável através das
opções anteriores (i.e., a postura ocorre como nas primeiras e segundas
escolhas nas latitudes mais elevadas e como na terceira e quarta nas latitudes
mais baixas).
·
Uma vez na vida (i.e.,
a postura ocorre apenas uma vez e é geralmente seguida da morte do indivíduo)
É de
notar que este campo se refere às espécies em geral e que a frequência de
postura pode ser diferente nas populações que se encontram no limite de
distribuição geográfica (latitude) da espécie.
O campo Quantidade De Postura indica se os
indivíduos têm uma postura múltipla durante a época de postura.
A classificação da estratégia de reprodução segue a
sugerida por E. Balon
A ESTRATÉGIA DE REPRODUÇÃO é
descrita na combinação de dois campos de escolha, seguido por uma classificação
sugerida por Balon (1990). O primeiro campo apresenta o tipo de cuidados
parentais com as escolhas: não
guardam; guardam; transportam. O segundo campo refere-se
ao padrão de protecção dos ovos ou juvenis, com as escolhas: postura em
substrato aberto (sem guardião, os ovos são deixados após a postura na coluna
de água ou no substrato, ex: rochas, gravilha, areia, plantas, etc.); postura
escondida (sem guardião, os ovos são colocados em locais imperceptíveis, ex.
grutas, fendas de rochas, depressões na areia, dentro de invertebrados vivos,
etc.); guardião de ovos (não fazem ninho, mas guardam os ovos à superfície, ou
por baixo de objectos ou nalgum substrato, ex. rochas, plantas, etc.); fazem
ninho (peixes que depositam e guardam
os ovos em ninhos, ex. bolas de muco, rochas, gravilha, areia, buracos, na base
de anémonas, plantas, etc,); portadores externos (peixes que incubam os ovos
externamente no corpo, ex. bolsas, boca, cavidades branqueais, barbatanas
pélvicas, etc.); portadores internos (fertilizam os ovos internamente, sendo o
desenvolvimento também interno, no corpo da mãe.)
Informação
sobre o ciclo de vida, acasalamento e comportamento de postura não incluídos
nesta lista de escolhas, e outras informações encontram-se no campo Descrição do Ciclo de Vida e Acasalamento.
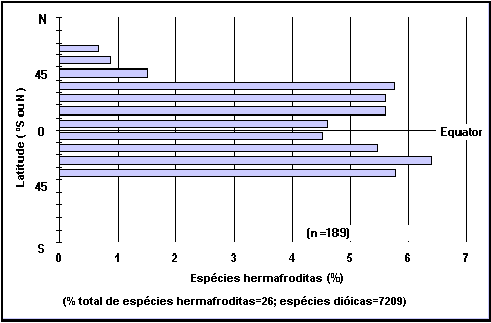
Caixa 20. A
distribuição de hermafroditismo por latitude.
A
forma mais comum de distribuição das gónadas em peixes é para as fêmeas
desenvolverem ovários e os machos testículos, e de assim funcionarem de modo
“dióico”.
Contudo,
em alguns grupos, indivíduos que incialmente eram fêmeas tornam-se machos ou o
contrário. É mais raro a ocorrência e funcionamento dos dois aparelhos
reprodutores em simultâneo no mesmo indivíduo (verdadeiro hermafroditismo),
apenas conhecido em Rivulus marmoratus.
As
espécies de peixes em que existe predominância das formas de hermafroditismo
mais comuns (o oposto de ocorrer em indivíduos isolados) representam apenas uma
pequena percentagem da totalidade das espécies de peixes, e estão concentradas
em famílias como Serranidae, Labridae, e Scaridae, e em baixas latitudes.
As
percentagens exibidas no gráfico de hermafroditismo por latitude da FishBase
não são tão precisas como seria desejável, uma vez que o gráfico pode ser
utilizado para gerar resultados assumindo apenas que que todas as espécies
actualmente sem entrada no campo “Modo” da tabela REPRODUÇÃO são dióicas. O
preenchimento deste campo para todas as espécies pode conduzir a alterações da
forma do gráfico, que apresenta no momento uma diminuição no Equador, e onde se
espera que esteja o máximo.
Daniel Pauly
Fig. 29. Percentagem de peixes
hermafroditas em relação à variação de latitude. Consulte a Caixa 20.
Estado Até à data mais de 700 registos de mais de 200
referências foram compilados. Mais de 50% das entradas foram verificadas na literatura
original, tal como Trewavas (1983) ou Balon (1990). Planeia-se aumentar
drasticamente a cobertura de formas de reprodução, utilizando os clássicos
Breder and Rosen (1966), Thresher (1984) e outras compilações.
Gráficos
Pode ser criado um gráfico de hermafroditismo vs latitude (veja a Caixa 20). Pode
aceder a este gráfico clicando consecutivamente nos seguintes botões: Relatórios no Menú Principal, Gráfico na janela Relatórios
Pré-definidos, Reprodução e Estados
Iniciais na janela Gráficos e Hermafroditismo
vs latitude.
Como Lá Chegar Chega-se à tabela REPRODUÇÃO clicando no botão Biologia na janela ESPÉCIES e no botão Reprodução na janela BIOLOGIA.
Referências Balon, E.K. 1990. Epigenesis of an epigeneticist: the
development of some alternative concepts on the early ontogeny and evolution of
fishes. Guelph Ichthyol. Rev. (1):1-48.
Breder, C.M., Jr. and D.E. Rosen. 1966. Modes of reproduction in fishes. T.F.H. Publications, Neptune City. 941 p.
Lagler, K.F., J.E. Bardach, R.R. Miller and D.R.
May-Passino. 1977. Ichthyology. Second edition. John Wiley and
Sons, New York. 506 p.
Thresher, R.E. 1984. Reproduction in reef fishes. T.F.H.
Publications, Neptune City. 399 p.
Trewavas, E. 1983. Tilapiine fishes of the genera Sarotherodon, Oreochromis and Danakilia.
British Museum of Natural History, London, UK. 583 p.
Armi Torres
Sobreviver
à maturação sexual e ser capaz de contribuir para o pool genético, define o
fitness de um individuo, e é crucial para a sobrevivência de uma população.
Para a gestão de um regime que assegure, na fase explorada, que um número
suficiente de juvenis chegue à primeira maturação, requer usualmente informação
sobre o tamanho e idade à qual o individuo atinge essa maturação.
É
conhecido que a maturidade sexual está associada a mudanças fisiológicas e
comportamentais, que mais tarde por vezes se manifestam sob a forma de
agregados de reprodutores, migração, territorialidade. A relação entre estas
mudanças biológicas e crescimento, mortalidade e longevidade foi estudada,
entre outros, por Beverton e Holt (1959), Alm (1959) e Pauly (1984) (veja a
Caixa 21).
Caixa 21 .
A capacidade reprodutora dos peixes.
Alguns
tópicos que parecem óbvios como a relação entre o crescimento e a reprodução
dos peixes, são no entanto mal compreendidos. O conhecimento convencional,
adoptado em inúmeros artigos, livros e relatórios, é de que os peixes tendem a
crescer mais rapidamente até atingirem a primeira maturação e, a partir dessa
altura, crescem mais devagar “porque a energia utilizada anteriormente no
crescimento somático (L¥) é agora utilizada para reprodução”. Esta hipótese pode ser denominada
por “Via reprodutora”.
Obviamente
que é provável que esta hipótese esteja errada e uma alternativa foi proposta:
é o abrandar do processo de crescimento que despoleta a maturação, em vez de ser
a maturação e desova a provocar a diminuição do crescimento (Iles 1974; Koch and Wieser 1983; Pauly 1984; Thorpe 1987).
Para
avaliar estas hipóteses, podemos examinar os seus corolários, ou seja, as
previsões esperadas. Sabe-se que muitos peixes pequenos crecem rapidamente até
um tamanho próximo de L¥ , e
depois desovam e reduzem o crescimento drasticamente. Pelo contrário, peixes de
grandes dimensões aproximam-se de L¥ gradualmente, com uma redução no crescimento perto
de ½ L¥ quando começam a desova. Este facto produz a
tendência de decrescente observada no gráfico de capacidade reprodutora vs comprimento assimptótico (Fig. 30),
que corrobora a segunda das hipóteses acima descritas. Por outro lado, a
primeira hipótese não explica um gráfico desta natureza, e a sua interpretação
iria necessitar outras hipóteses ad hoc.
Referências
Iles, D. 1974. The tactics and strategy of growth in fishes, p. 331-345. In F.R. Harden Jones (ed.) Sea
fisheries research. Elek Science, London.
Koch, F. and W. Wieser. 1983. Partitioning of
energy in fish: can reduction in swimming activity compensate for the cost of
production? J. Exp. Biol. 107:141-146.
Pauly, D. 1984. A mechanism for the juvenile-to-adult transition in fishes.
J. Cons. CIEM 41:280-284.
Thorpe, J.E. 1987. Smolting versus
residency: developmental conflict in salmonids. Am. Fish. Soc. Symp. 1:244-252.
Daniel Pauly
Existem registos de maturação de mais de 800 espécies
provenientes de mais de 400 referências
Fontes A tabela MATURAÇÃO contém informação do comprimento e
idade da primeira maturação sexual de mais de 800 espécies (mais de 1800
registos) de mais de 400 referências. Entre as maiores fontes de informação
encontra-se Beverton e Holt (1959), Compagno (1984a, 1984b), Dorel (1985) e van
der Elst and Adkin (1991).
Na
literatura, a informação sobre a maturação sexual aparece sob a forma de várias
categorias relacionadas entre si:
1.
moda ou comprimento
médio, ou idade, i.e., O comprimento ou idade ao qual 50% da população atinge a
primeira maturação sexual.
2.
o comprimento ou idade
ao qual, uma certa percentagem (não 50%) da população atinge a maturação.
3.
o comprimento ou idade
do menor peixe maturo
4.
o comprimento ou idade
do maior peixe que atinge a 1ª maturação sexual.
5.
como intervalo de
comprimento ou idade do menor (mais novo) ao maior (mais velho) peixe maturo (3
e 4)
6.
como intervalo de
tamanhos e idades médios de maturação
7.
valores não
qualificados
Inicialmente esta tabela incluía apenas informação
sobre a moda e comprimento médio ou idade (categoria 1). Esses valores eram
usualmente derivados de uma interpolação linear, análise de probabilidades,
pelo ajustamento da curva logística, ou então através de um conjunto estado de
maturação/comprimento ou idade. No entanto, na maior parte dos casos, o método
utilizado não é mencionado. A tabela foi sendo modificada para acomodar as
diferentes informações existentes.
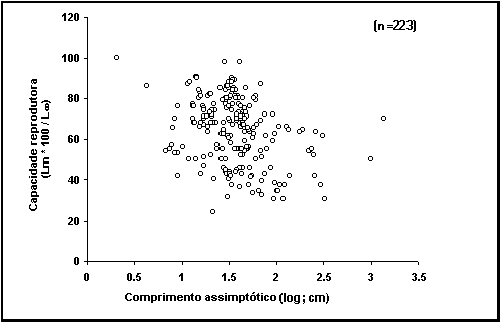
Fig. 30. Capacidade reprodutora para
vários peixes. Repare na tendência decrescente e veja a Caixa 21.
Campos A 1ª categoria entra nos campos Lm e tm.
Os campos de intervalo acomodam a
informação das categorias 2 a 7. Valores não qualificados (categoria 7) entram
como valores mínimos no campo intervalo.
Nalguns casos, o campo Comentários,
esclarece algum valor entrado.
Estado Para verificar o comprimento da 1ª maturação sexual (Lm), verifica-se se a razão
correspondente L¥/Lm se mantém dentro do intervalo conhecido para os
peixes (Beverton e Holt, 1959).
Relatórios Pode acerder a um relatório prédefinido através do
Menu Principal da FishBase, com informação por família desta tabela, ou se
clicar no ícone do gráfico pode ver o gráfico capacidade reprodutora vs comprimento assimptótico. Este
gráfico pode também ser acedido a partir da tabela MATURAÇÃO.
Como Lá Chegar Chega-se à tabela MATURAÇÃO clicando no botão Biologia na janela ESPÉCIES, no botão Reprodução na janela BIOLOGIA, e no
botão Maturação na janela REPRODUÇÃO
E ESTADOS INICIAIS. Chega-se ao relatório da Maturação, clicando no botão Relatórios no Menu principal, no botão Dinâmica Populacional Por Família na
janela RELATÓRIOS PRÉ-DEFINIDOS e no botão Informação
Sobre Maturação na janela seguinte.
Referências
Alm, G. 1959. Connection between maturity, size, and age in fishes. Inst. Freshwat. Res. Rep.
No. 40. 145 p.
Beverton, R.J.H. and S.J. Holt. 1959. A review of the lifespans and mortality rates of fish in
nature, and their relation to growth and other physiological characteristics,
p. 142-180. In G.E. Wohstenholme and
M. O’Conner (eds.) CIBA foundation colloquia on ageing. Vol. 5. The lifespan of
animals. J. and A. Churchill, Ltd., London. 344 p.
Compagno, L.J.V. 1984a. FAO species catalogue. Vol. 4. Sharks of the world. An
annotated and illustrated catalogue of shark species known to date. Part 2.
Carchariniformes. FAO Fish. Synop. 4(125)Pt. 2:655 p.
Compagno, L.J.V. 1984b. FAO species catalogue. Vol. 4. Sharks of the world. An
annotated and illustrated catalogue of shark species known to date. Part 1.
Hexanchiformes to Lemniformes. FAO Fish. Synop. 4(125) Pt. 1, 249 p.
Dorel, D. 1985. Poissons de l’Atlantique nord-est:
relations taille-poids. Institut Français de Recherche pour l’Exploration de la
Mer. 165 p.
Pauly, D. 1984. A mechanism for the juvenile-to-adults
transition in fishes. J. Cons. CIEM 41:280-284.
van der Elst, R.P. and F. Adkin, Editors. 1991. Marine linefish: priority species
and research objectives in southern Africa. Oceanogr. Res. Inst. Spec. Publ.
No. 1, 132 p.
Crispina Binohlan
Existe uma postura da população de arenque em cada
época do ano
As épocas de postura podem
variar consideravelmente entre populações da mesma espécie. Por exemplo, no
Atlântico Norte, existe uma postura de uma população de arenque em cada época
do ano.
A tabela
POSTURA apresenta informação sobre a época de postura, razão entre os sexos,
fecundidade absoluta e relativa, relação entre fecundidade e comprimento,
frequência diária de postura dos diferentes stocks (populações) da mesma
espécie em diferentes localidades.
Campos Os campos País
e Localidade identificam os locais
de postura, enquanto que o Local de
Postura refere-se ao tipo de habitat onde a postura ocorre, o qual pode
ser: lacustre, fluvial, estuarina, costeira, na plataforma continental ou
oceânica.
A Época de postura define o mês do ano na
qual tem lugar a postura. A % mensal de fêmeas maduras pode ser aqui
encontrada. Quando é usado 111,
refere-se ao mês na qual as fêmeas maduras foram encontradas, mas sem indicação
da sua abundância relativa.
Pode ser
criado um gráfico que mostra a época da reprodução para um dado stock, baseado
em percentagens ou entradas de valores 111. No último caso, os dados são
integrados (mais de três meses), o que dá origem a erros padrão aproximados.
Além disto, também pode ser criado um gráfico composto que combina dados de
vários gráficos num único plot. Quando os respectivos erros padrão são baixos,
isto indica uma época de postura
similar para todos os stocks de uma mesma espécie (Fig. 31).
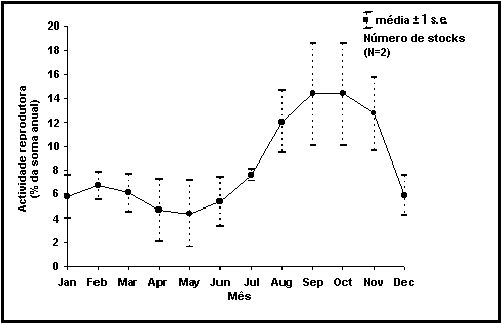
Fig. 31. Época de postura de Engraulis ringens a Norte /Centro do Perú
O campo Temperatura, dá o intervalo de
temperaturas da água em graus Celsius, no qual geralmente a postura ocorre,
enquanto que o campo razão entre os
sexos (%) mostra a % média de fêmeas em postura no stock em postura. Se uma
razão entre os sexos (%) publicada
for dada como fracção (nº de fêmeas/nº de machos) então foi transformada
usando:
A fertilidade pode ir de 300 milhões de ovos até a
uma pequena descendência
![]()
Fecundidade
define-se como o nº de óvulos encontrados numa fêmea em postura, e é
frequentemente encontrado na literatura sem indicação do peso do corpo
correspondente. Apesar desta informação ser menos útil que a fecundidade
relativa (ver abaixo), foi decidido incluí-la nesta tabela. Para incluir casos
em que a informação tamanho está disponível, adicionou-se campos que permitem a
entrada, para cada registo de fecundidade, um intervalo de pesos e tamanhos dos
indivíduos. Foi também providenciado um campo de escolha para identificar o
tipo de medida utilizado. As escolhas consistem em: SL (comprimento standard); FL
(comprimento do corpo; TL
(comprimento total); WD (largura do
disco dos raios); NG ( não dado na fonte) e OT (outros comprimentos).
A relação fecundidade/comprimento é raramente
encontrada na literatura
A Fecundidade Relativa é dada quando disponível e define-se como o
número de oócitos maduros na fêmea, dividido pelo peso total da mesma.
A
relação Fecundidade/Comprimento é
uma informação útil, mas raramente encontrada na literatura. No entanto, foram
providenciados campos de entrada com esta informação, da seguinte maneira:
Tamanho:
são dois campos que referem o maior e menor peixe considerado, quando a relação
fecundidade/ comprimento foi estabelecida. O campo de escolha define o tipo de
comprimento utilizado (ver acima).
a:
refere-se ao factor multiplicativo (a) da relação fecundidade/comprimento sob a
forma de F= a Lb, onde F é a fecundidade em número de ovos, e L é o comprimento em centímetros.
b:
refere-se ao expoente da relação fecundidade/ comprimento.
n:
refere-se ao número total de espécimes utilizados para derivar a relação
fecundidade/comprimento
r: é o
coeficiente de correlação da relação fecundidade/ comprimento na sua forma log-linear.
A Frequência Diária de Postura aplica-se
apenas a posturas em grupo e dá a frequência de postura por dia (ex. 0,5 indica
que metade das fêmeas fazem a postura todos os dias, i.e., um indivíduo fêmea
faz a postura todo o segundo dia, ver Hunter e Goldberg 1980, Hunter e Leong
1981 ou Pauly e Soriano 1987.
Informação
adicional sobre os locais e época de postura é dado no campo Comentários.
Status A tabela
POSTURA contém mais de 1400 registos de quase 1000 espécies. Muitas entradas
apenas têm a época de reprodução, mas cerca de 500 têm também a razão entre os sexos ou a fecundidade.
Esta informação foi extraída de mais de 300 referências, das quais se salienta
Allen (1985), Collette and Nauen (1983), van der Elst and Adkin (1991) e Koli
(1990).
Mais
de 50% da informação desta tabela foi verificada. A cobertura actual expandirá,
e assimilará gradualmente o enorme volume de literatura disponível sobre
postura, especialmente nas espécies comerciais.
Como Lá Chegar Chega-se à tabela POSTURA clicando no botão Biologia na janela ESPÉCIES, no botão Reprodução na janela BIOLOGIA, e no
botão Postura na janela seguinte.
Referências Allen, G.R. 1985. FAO species catalogue. Vol. 6.
Snappers of the world. An annotated and illustrated catalogue of lutjanid
species known to date. FAO Fish. Synop. 6(125):208 p.
Collette, B.B. and C.E.
Nauen. 1983. FAO species catalogue. Vol. 2. Scombrids of the world.
An annotated and illustrated catalogue of tunas, mackerels, bonitos and related
species known to date. FAO Fish. Synop. 2(125):137 p.
Hunter, J.R. and S.R. Goldberg. 1980. Spawning incidence and batch fecundity in northern anchovy, Engraulis mordax. U.S. Fish. Bull. 77:641-652.
Hunter, J.R. and R. Leong. 1981. The spawning energetics of female northern anchovy, Engraulis mordax. U.S. Fish. Bull.
79:215-230.
Koli, L. 1990. Suomen kalat. [Fishes of Finland]. Werner
Söderström Osakeyhtiö, Helsinki. 357 p.
Pauly, D. and M. Soriano. 1987. Monthly spawning stock and egg production of Peruvian anchoveta (Engraulis ringens), 1953 to 1982, p.
167-178. In D. Pauly and I. Tsukayama
(eds.) The Peruvian anchoveta and its upwelling ecosystem: three decades of
change. ICLARM Stud. Rev. 15, 351 p.
van der Elst, R.P. and F. Adkin, Editors. 1991. Marine linefish: priority species
and research objectives in southern Africa. Oceanogr. Res. Inst. Spec. Publ.
No. 1, 132 p.
Armi Torres